Chapitre 5 : Transport et stockage de l’oxygène
1 Hémoglobine et Myoglobine
L’Oxygène est un thème récurrent à cause de son rôle dans la respiration et sa production dans la photosynthèse. Ces deux cas fournissent des exemples d’une gamme de réactions rédox, de transferts d’électrons, de transferts d’atome, et de processus photochimiques biologiquement importants. Pour assurer un approvisionnement en O2, trois types de protéines sont concernés, ils lient et transportent l’oxygène. Il s’agit de la famille des hémoglobines-myoglobines, les hémocyanines et hémérythrines. Ils sont appelés communément les transporteurs d’oxygène. Chacune de ces protéines utilise un atome métallique, ou une paire d’atomes métalliques dans des protéines qui se sont développées pour être utilisées par un type d’organisme particulier. Dans ces protéines, il y a un équilibre fragile dans lequel la molécule d’O2 se lie à un centre contenant du fer ou du cuivre, sans subir de transfert électronique irréversible, ni de réaction d’oxydo-réduction conduisant au clivage de la liaison O—O et aux réactions d’oxydation concomitantes.
Le transporteur et distributeur d’oxygène le plus connu et le plus répandu est l’hémoglobine (figure 1).
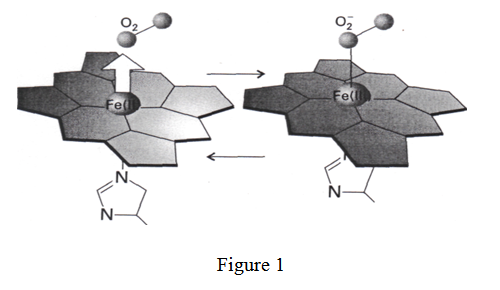
L’hémoglobine est présente dans les globules rouges du sang. Il est utilisé par les vertébrés pour transporter l’oxygène des poumons ou des branchies vers les tissus où l’O2 doit être utilisé dans un processus qui le réduit en CO2. Les vertébrés stockent l’oxygène dans la myoglobine. Cette molécule a un site coordinant de l’O2 semblable à celui de l’hémoglobine. La myoglobine fournit de l’oxygène aux muscles pour entretenir l’effort. Dans l’hémoglobine (Hb) et la myoglobine (Mb), le site de liaison de l’oxygène est un complexe fer-porphirine qui subit un changement de structure à la suite de la liaison d’O2.
Un second type de transport d’oxygène est l’hémocyanine (figure 2). On le trouve dans les escargots (mollusques) et les crabes (arthropodes).
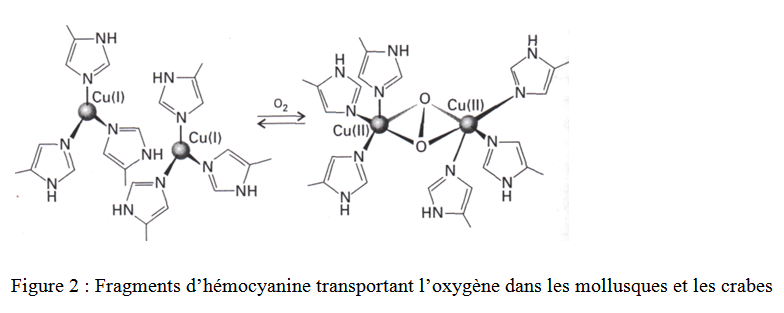
L’O2 est lié à deux ions Cu+ reliés directement à la protéine. Les interactions entre l’O2 et les sites métalliques transforment les Cu+ en Cu2+ et l’O2 en O22-.
Le troisième type de transporteur d’Oxygène est l’hémérythrine (figure 3).
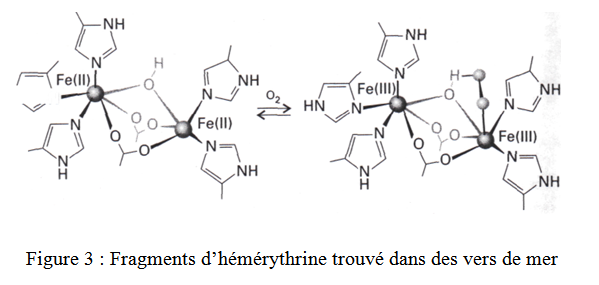
On le trouve dans certains vers marins. Le site de fixation de l’oxygène de l’hémérythrine est constitué de deux ions ferreux voisins. Ils sont directement liés aux atomes donneurs de la protéine plutôt que sur le macrocycle de l’Hème. L’O2 est lié à un des Fe comme un hydroperoxyde (HO2-). Curieusement, certains organismes qui utilisent l’hémérythrine pour transporter l’oxygène, se servent d’une protéine totalement différente, la myoglobine, pour stocker l’Oxygène.
O2 se fixe à une porphyrine de fer dans l’hémoglobine et la myoglobine, à une paire d’ions cuivre liés à la protéine dans l’hémocyanine et à l’un des deux atomes de fer liés à la protéine dans l’hémérythrine.
La protéine hélicoïdale de l’hémoglobine (Hb) joue un rôle régulateur intéressant dans le fonctionnement de ce transporteur d’oxygène. Les spires de cette protéine agissent comme des ressorts qui peuvent répondre à la tension générée lorsque l’oxygène se fixe à l’une des porphyrines de fer et que la protéine transmet cette tension aux autres sites. Cette interaction a pour résultat d’augmenter l’affinité pour O2 du deuxième site. Ce phénomène, appelé coopérativité, permet à l’hémoglobine de fixer et de libérer O2 plus facilement. Nous voyons que la fixation de O2 peut être une combinaison de la complexation sur le site métallique et de l’altération de l’environnement protéinique.
L’hémoglobine se trouve essentiellement à l’intérieur des globules rouges du sang (13,5 à 17,5 gr/dL chez l’homme et 12,5 à 15,5 gr/dL chez la femme). L’Hb humaine est constituée de 4 chaînes identiques deux à deux : deux chaînes de α de 141 acides aminées chacune et deux chaînes de β acides aminées chacune. Chacune de ces chaînes est associée à un groupement prosthétique : l’héme.
L’O2 se comporte comme un π accepteur puissant dans son interaction avec le Fe2+. Dés lors, il n’est pas surprenant que d’autres ligands π accepteurs soient capables de se lier au fer de l’hémoglobine ou de la myoglobine. Les complexes avec le NO, CO, CN-, RCN, N3- et SCN- ont été étudiés. Ces ligands fortement liants peuvent totalement altérer la fonction de l’hémoglobine, allant jusqu’à causer un résultat fatal.
Les polypeptides jouent un rôle important dans l’hémoglobine. Le complexe de la porphyrine du fer sans les polypeptides est oxydé irréversiblement en Fe3+ par l’O2 formant un μO2- stable qui ne peut pas fonctionner comme un transporteur d’O2.
Cette dimérisation oxydante improductive des porphyrines de fer est potentiellement un défaut fatal pour leur fonction biologique. Toutefois, elle est protégée dans l’hémoglobine ou la myoglobine par les chaînes polypeptidiques qui entourent et protègent le site Heme.
La structure du dioxymyiglobine, la forme libre de l’oxygène dans la myoglobine, est illustrée sur la figure 4.
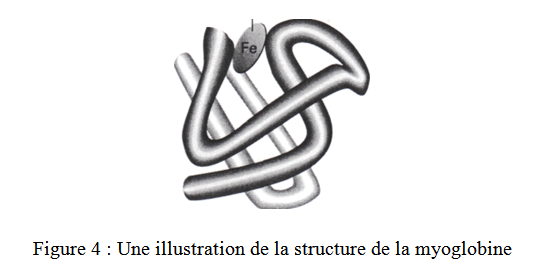
Il faut remarquer que le site actif de la porphyrine est entrelacé par la chaîne polypeptidique. La poche de protéine dans laquelle l’hème est piégé, est formée par les résidus d’aminoacides ayant des groupements latéraux apolaires qui la rendent hydrophobe. Ces mêmes groupements bloquent l’accès des molécules plus grandes au Fe et ainsi la formation d’une espèce pontée Fe—O2---Fe.
Les groupements hydrophobes empêchent aussi la solvatation des ions produits lors de l’oxydation du complexe du fer. Le résultat étant que le complexe du Fe2+ puisse survivre assez longtemps pour lier et libérer l’O2. Ceci est une illustration typique du contrôle délicat de l’environnement de la réaction que la protéine peut exercer.
Le déoxy Mb est un complexe pentacoordiné du Fe2+ High spin avec 4 des positions de coordination occupés par les N de la porphyrine et la 5éme position est occupée par un N de l’imidazole du résidu histidine qui couple l’heme à la protéine (figure 5).
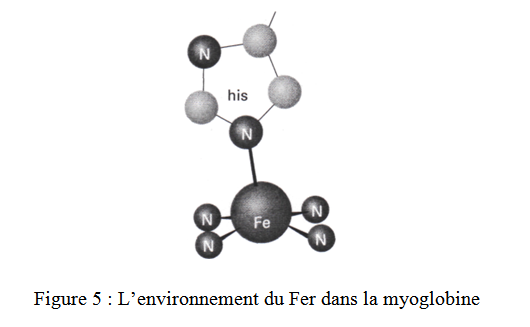
Cette pentatcoordination de l’heme du complexe du Fe2+ est toujours High spin t2g4eg2. L’ion ferreux high spin est plus grand que la cavité de la porphyrine et ainsi se met à 0,4Å au dessus du plan de la porphyrine (4).
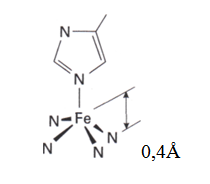
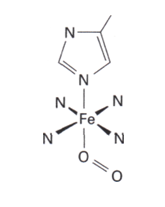
Dans les complexes porphyrines du fer(II) High spin, tels que l’hémoglobine et la myoglobine, le macrocycle de la porphyrine est plissé et tordu, alors que l’ion Fe(II) low spin qui est plus petit peut s’insérer confortablement dans le cycle.
Quand le O2 complète la sixième coordination, le complexe devient low spin (t2g6) et l’ion Fe2+ se contracte un peu et descend dans le trou du plan. Ce changement structurel a été clairement mis en évidence par la diffraction des rayons X.
Les protéines hélicoïdales associées à l’hémoglobine et la myoglobine facilitent la fixation coopérative de O2 et empêchent la formation de dimères oxo non fonctionnels.