Chapitre 5 : Transport et stockage de l’oxygène
4 Coopérativité
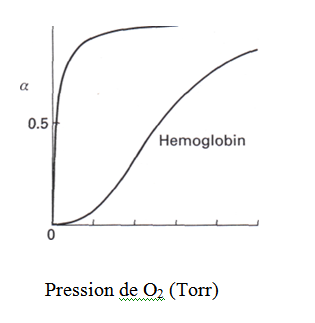
La fonction de l’hémoglobine consiste à fixer O2 aux pressions partielles élevées du tissu pulmonaire, à le transporter sans perte par le sang et à le libérer au profit de la myoglobine des tissus cellulaires. Ceci requiert que le Mb ait une plus grande affinité à l’O2 que Hg aux faibles pressions partielles. C’est le cas, voir figure 7.
Figure 7 Les courbes de saturation d’Oxygène de Mb et Hg montrant la fraction de saturation α en fonction de la pression partielle d’oxygène à pH = 7,2.
La forme de la courbe de Mb dans la figure peut être représentée par un équilibre simple :
Mb + O2 ----------> MbO2
[MbO2]
K = ---------------
[Mb]p
p est la pression partielle de O2.
[MbO2]
La fraction de saturation d’oxygène α est : α = ---------------------------
[Mb] + [MbO2]
Il vient α = kp/1+Kp
La courbe du Mb est une illustration parfaite de cette équation mais la courbe de Hg ne l’est pas. La courbe du Hg est décrite par une fonction plus complexe où la dépendance de α sur la pression partielle de l’oxygène est remplacé par pn avec n compris entre 2 ou 3.
Le pH est aussi un facteur important dans la formation de la liaison entre l’oxygène et l’Hg bien qu’il n’y a pas de groupements acides sur l’heme. Il est observé que l’Hg relâche plus facilement l’O2 quand le pH est faible. Il vient dés lors que l’O2 est libéré plus aisément dans les cellules ou le métabolisme est actif résultant d’une concentration élevé de CO2 avec des pH plus faibles. Un autre détail à considérer, les phosphates organiques de la protéine de l’Hg bien qu’éloignés de l’hème, affecte la liaison de l’hème avec l’oxygène.
La différence structurale entre l’Mb et l’Hg provient de ce que Hb est essentiellement un tétramère de Mb contenant quatre groupements hème (fig I.9), alors que Mb n’en contient qu’un. Cette différence est crucial parce qu’elle permet aux quatre unités hèmes de Hb de lier l’O2 de façon coopérative : une fois qu’un O2 est fixé à Hb, l’affinité pour d’autres est augmentée.

Lorsque Fe(II) High spin, qui se trouve à 0,4Å au-dessus du plan de la porphyrine, coordine O2, il devient low spin et se déplace jusqu’au plan en entrainant avec lui le résidu histidine de la protéine. Il en résulte que la forme de la protéine change et que les caractéristiques de fixation des autres sites sont modifiées. On attribue la dépendance par rapport au pH et au phosphate à des influences conformationnelles provenant de points modérément distants de l’atome métallique. Ce dernier changement souligne une fois de plus la sensibilité des structures à des influences sur des sites éloignés.
Le phénomène de la coopérativité promeut le transfert de l’O2 de la protéine de prélèvement, l’hémoglobine (Hg,) à la protéine de stockage, la myoglobine (Mg).