Chapitre 4 : Les éléments dans les milieux vivants
Chapitre 4 : Les éléments dans les milieux vivants
| Site: | Touch By SukaJanda01 |
| Cours: | CHIMI 5121 : Chimie bio inorganique |
| Livre: | Chapitre 4 : Les éléments dans les milieux vivants |
| Imprimé par: | Visiteur anonyme |
| Date: | mardi 17 juin 2025, 13:30 |
1 Éléments chimiques essentiels
Les organismes vivants n’utilisent pas les éléments de la même façon que leur ordre d’importance sur terre (tableau1)
Tableau 1 : Abondance en masse des éléments par ordre décroissant
|
Terre Ecorce |
Océan |
Êtres Humains |
|
O |
O |
Oxygène (53,6%) |
|
Si |
H |
Carbone (16%) |
|
Al |
Cl |
Hydrogène (13,4%) |
|
Fe |
Na |
Azote (2,4%) |
|
Ca |
Mg |
Sodium, potassium, Sulfure (0,1%) |
|
Mg |
S |
Chlore (0,09%) |
|
Na |
Ca |
Silicium, magnésium (0,04%) |
|
K |
K |
Fer, Fluor (0,005%) |
|
Ti |
C |
Zinc (0,0003) |
|
H |
Br |
Cuivre, Brome (2.10-4%) |
|
P |
B |
Selénium, Manganése, Arsenic, Nickel (2.10 –5%) |
|
|
|
Plomb, Cobalt (9.10-6%) |
Le tableau 1 montre une abondance de l’aluminium et du Silicium dans l’écorce terrestre alors que ces éléments ne sont que faiblement présents chez les mammifères. L’ordre d’importance dans la nature et les océans n’est pas celui des mammifères. Quelques éléments qui sont présents à l’état de trace dans la croûte terrestre, jouent un rôle central dans les organismes contribuant aux équilibres dynamiques : le Cuivre et le Sélénium en sont des exemples.
Les éléments chimiques dans le système biologique peuvent être classés en quatre groupes :
- Les éléments en grande proportion : H/H+, C, N, O2-/O2-/O22-, P, S/S2- ;
- Les macro-minerais et ions : Na/Na+, K/K+, Mg/Mg2+, Ca/Ca2+, Cl-, PO42-, SO42- ;
- Les éléments sous forme de trace : Fe/Fe2+/Fe3+/Fe4+, Zn/Zn2+, Cu/Cu+/Cu2+/Cu3+ ;
- Les éléments présents sous forme d’ultra trace qu’on peut classer en :
- Non métaux : F/F-, I/I-, Se/Se2-, Si/Si4+, As, B ;
- Mn/Mn2+/Mn3+/Mn4+, Mo/Mo4+/Mo5+/Mo6+, Co/Co2+/Co3+, Cr/Cr3+/Cr5+, V/V3+/V4+/V5+, N+/Ni3+, Cd/Cd2+, Sn/Sn2+/Sn4+, Pb/Pb2+, Li/Li+.
L’essentiel des éléments en biologie relève de certains critères :
- Une déficience physiologique apparaît quand l’élément est enlevé par un régime ;
- La déficience est corrigée par l’addition de l’élément ;
- Une fonction biologique spécifique est associée à l’élément.
Chaque élément essentiel requiert une courbe de dosage (figure 1). Aux faibles concentrations, l’organisme ne survie pas tandis que dans la région de déficience, l’organisme ne donne pas la réponse optimale. Nous avons après le plateau de concentration optimale et aux plus grandes concentrations, l’élément devient toxique pouvant même conduire à la mort.
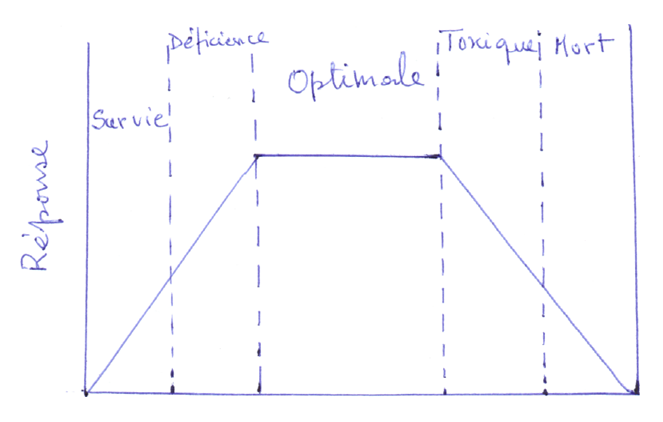
Dosage des éléments
μg/l 10 50 Se 200 103 104
mg/l 0,5 2 F 10 20 100
En plus du facteur d’existence, beaucoup de propriétés chimique et physique des éléments et ions sont responsables des processus dans les systèmes biologiques. Cela dépend des charges ioniques, des rayons ioniques, de la sélectivité des ligands, de la préférence de la coordination et de la géométrie, du couplage de spin, du contrôle cinétique systémique, de la réactivité chimique des ions en solution.
2 Rôle des ions métalliques en biologie
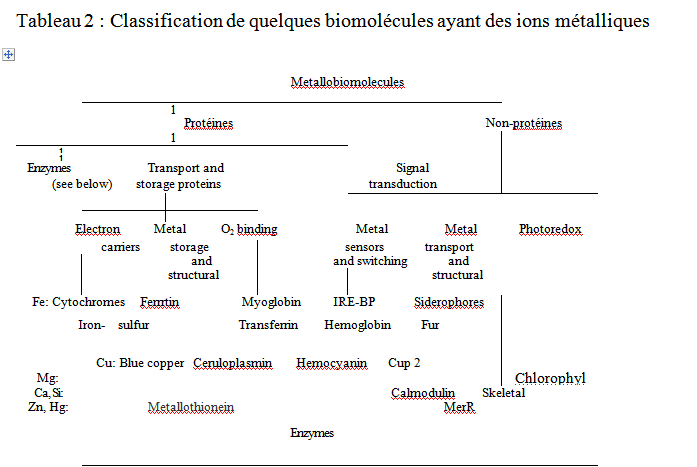
Le tableau 2 résume les biomolécules qui utilisent des ions métalliques. Beaucoup de ces molécules sont des protéines. Environ 30% des enzymes ont un ion métallique sur leur site actif. Ces métallo-enzymes facilitent des réactions variées, comme l’hydrolyse acido-catalysée (réalisée par les hydrolases), les réactions rédox (oxydases et oxygénases) et les réarrangements des liaisons carbone carbone (synthases et isoméras). Les ions métalliques ont aussi des rôles structuraux. Par exemple, l’ion Ca2+ est impliqué dans les déformations des protéines. Le tableau 3 montre que les molécules contenant des ions métalliques remplissent des fonctions importantes comme le transport d’électrons, de sites de stockage des métaux, de sites de liaison et de stockage d’Oxygène, et la transmission de signaux. Les ions métalliques sont aussi impliqués dans des réactions photo-redox, comme c’est le cas pour la chlorophylle, une molécule qui capte de l’énergie.
Bien que certaines fonctions mentionnées soient partagées avec des espèces non métalliques, il est clair que les complexes métalliques jouent un rôle majeur en biologie. Leurs modes d’interventions sont diverses et variées. Les ions métalliques jouent un effet inductif par la coordination au site de réaction, servent de sites rédox qui fonctionnent par transfert d’électron ou d’atome. La sélectivité s’opère par le biais d’ions ayant la taille approprié, la préférence stéréochimique, le caractère soft-hard, ou le potentiel réducteur.
Tableau 2 : Classification de quelques biomolécules ayant des ions métalliques

Nous traiterons dans une large mesure des polypeptides. Ces macromolécules sont formées à partir de α aminoacide liés par une liaison peptidique.
H(CHR1R2)N ----C(O)—C(HRN)
Les vingt différents acides α aminés présents naturellement dans les protéines sont listés dans le tableau 3 avec leurs abréviations.
Notons que les chaines de ces aminoacides ont divers groupes fonctionnels : alkyle, carboxyle (COOH), amino (--NH2), hydroxyle (--OH) et thiol (--SH). Les groupements alkyls confèrent un caractère hydrophobe et les groupements plus polaires confèrent un caractère plus ou moins hydrophile. Beaucoup de ces groupements polaires servent d’acides au sens de Bronsted, ou de bases au sens de Lewis dans leur complexation avec des ions métalliques. Un rôle important des résidus peptidiques est la modification de l’environnement immédiat des ions métalliques dans les métallo-enzymes (tableau 3).

En plus de leur rôle dans les processus biologiques, les ions métalliques, sous forme de minerai cristalline ou de solides amorphes, sont importants comme des matériaux de structure de nombreux organismes (tableau 4).
Tableau 4 : Minerais dans des structures biologiques
|
Minerai |
Formule |
Organisme |
Localisation |
|
Calcite |
CaCO3 |
Oiseaux |
Coquille des œufs |
|
Aragonite |
CaCO3 |
Mollusques |
Coquilles |
|
Hydroxyapatite |
Ca5(PO4)3(OH) |
Vertébrés Mammifères |
Os |
|
Dioxyde de Silicone |
SiO2nH2O |
Diatomés Limpets Plantes |
Cellule de protection Dents Feuilles |
La principale matière première pour la production du phosphore élémentaire et de l’acide phosphorique est constituée par les roches phosphatées, les restes insolubles, écrasés et compactés d’organismes anciens, dont les minéraux principaux sont la fluoroapatite Ca5(PO4)3F et l’ hydroxyapatite Ca5(PO4)3OH.
Ca5(PO4)3F(s) + 5H2SO4(I) → 3H3PO4(I) + 5CaSO4(s) + HF (aq)
Le polluant potentiel, le fluorure d’hydrogène, qui vient du fluorure de la roche, est piégé par une réaction avec les silicates pour donner l’ion complexe moins réactif [SiF6]2-
L’hydroxyapatite est largement répandue dans le milieu animal comme le constituant principal des os, des dents et des coquilles. Dans les minéraux il existe avec d’autres substances, par exemple dans les os et les dents, l’hydroxyapatite coexiste avec la protéine collagène qui contrôle sa morphologie, son développement et influence ses propriétés mécaniques. Le Silicium est moins répandu dans les composés de la nature. Une exception est à noter, l’utilisation majeure de l’hydroxyle de Silicium dioxyde dans les diatomées qui vivent dans les mers.
En résumé, les ions métalliques sont utilisés pour la catalyse, le signalement, le stockage d’énergie, la maintenance des structures soft-hard des matériaux biologiques.
3 Biochimie du calcium
Bien qu’existant dans les biomatériaux durs, le Ca2+ a plusieurs bio-fonctions. Il sert de messager dans l’action hormonale, de déclencheur de la contraction musculaire, intervient dans l’amorçage de la coagulation du sang et dans la stabilisation de la structure des protéines.
La diffraction des rayons X et les résultats NMR montrent comment ces fonctions sont contrôlées par les changements de conformations induits par le Ca2+ quand il est lié au Calmodulin troponin C et des protéines similaires. Ces protéines sont impliquées dans l’activation des canaux et des récepteurs membranaires à la surface des cellules.
Les nombreux rôles du Ca2+ partent de son affinité avec les ligands oxygénés réputés hards en conjonction avec la labilité intermédiaire de ces complexes, qui se situe entre celle des ions alcalins et les ions des métaux d d’une part, et celle des ions moins labiles de ses congénères plus légers du groupe 2 (Be2+ et le Mg2+) d’autre part.
Les différences entre le Ca2+ et le Mg2+ sont révélatrices et sont fonctions de trois facteurs :
a) Comme conséquence de sa faible sélectivité, Ca2+ peut se lier aux oxygènes neutres des ligands donneur (carbonyles, et alcools) en compétition avec l’eau ;
b) Ca2+ ressemble au Na+ et K+ en favorisant un nombre de coordination élevé et une géométrie irrégulière. Avec un nombre de charge de +2, Ca2+ peut être lié à des anions que les ions alcalins ne peuvent pas ;
c) Les vitesses de fixation et de dissociation sur Ca2+ sont élevées : la vitesse de fixation est limitée par la diffusion et la vitesse de dissociation correspond à la stabilité. La vitesse d’exchange sur Mg2+ est
105s-1, plus lente que celle Ca2+de presque trois ordres de grandeur. Cette vitesse d’exchange rapide, qui est celle d’un système très réactif, explique l’utilisation du Ca2+ quand des réponses rapides sont nécessaires comme l’ouverture et la fermeture des canaux membranaires, pour la régulation du transport ionique et la contraction musculaire.
Les protéines qui fixent le calcium sont en général riches en aspartate et en glutamate, qui portent tous deux des groupements carboxylate sur leur chaîne latérale et qui peuvent donc agir comme ligands anioniques durs. Comme illustré dans une forme de structure qui existe, un Ca2+ entouré de 4 carboxylate avec des oxygènes donneurs, la géométrie est un tétraèdre déformé (figure 2). La structure a été caractérisée par diffraction des rayons X.
L’ion Ca2+ fonctionne communément comme un pont entre différents segments de la protéine. Il est lié aux groupements latéraux anioniques d’aminoacides différents, ou même à des groupements carbonyle (figure 2). L’utilisation de cette propriété dans le contrôle du repliement de la chaine est illustrée schématiquement sur la figure 2 qui montre comment la chaine peptidique se replie pour permettre aux quatre COO- de se coordiner au Ca2+. Le résultat de ce mouvement est le déplacement des emplacements des groupements latéraux de l’une des régions hélicoïdales de la protéine par rapport aux groupements latéraux de la deuxième région hélicoïdale.
Les variations du repliement des protéines contrôlent la structure et les fonctions de la cellule telles que sa vitesse de croissance et son métabolisme énergétique.
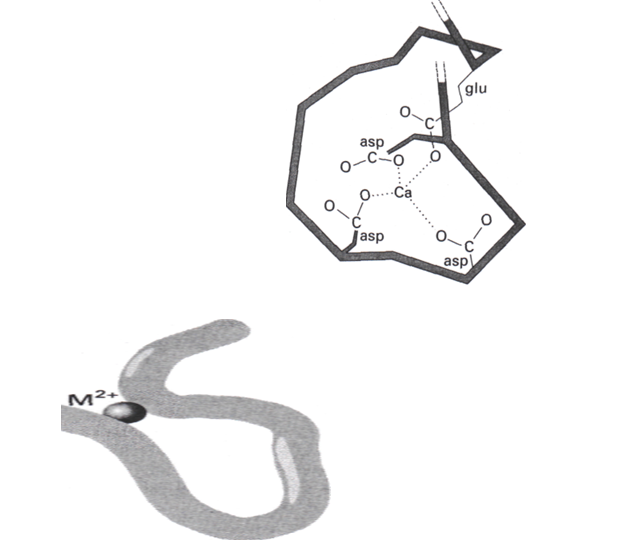
Figure 2
La position des ions Ca2+ et Mg2+ influence les déformations des protéines et par conséquent change les structures et fonctions des cellules, la labilité du Ca2+ permet les changements structuraux rapides.