Chapitre 4 : Les éléments dans les milieux vivants
3 Biochimie du calcium
Bien qu’existant dans les biomatériaux durs, le Ca2+ a plusieurs bio-fonctions. Il sert de messager dans l’action hormonale, de déclencheur de la contraction musculaire, intervient dans l’amorçage de la coagulation du sang et dans la stabilisation de la structure des protéines.
La diffraction des rayons X et les résultats NMR montrent comment ces fonctions sont contrôlées par les changements de conformations induits par le Ca2+ quand il est lié au Calmodulin troponin C et des protéines similaires. Ces protéines sont impliquées dans l’activation des canaux et des récepteurs membranaires à la surface des cellules.
Les nombreux rôles du Ca2+ partent de son affinité avec les ligands oxygénés réputés hards en conjonction avec la labilité intermédiaire de ces complexes, qui se situe entre celle des ions alcalins et les ions des métaux d d’une part, et celle des ions moins labiles de ses congénères plus légers du groupe 2 (Be2+ et le Mg2+) d’autre part.
Les différences entre le Ca2+ et le Mg2+ sont révélatrices et sont fonctions de trois facteurs :
a) Comme conséquence de sa faible sélectivité, Ca2+ peut se lier aux oxygènes neutres des ligands donneur (carbonyles, et alcools) en compétition avec l’eau ;
b) Ca2+ ressemble au Na+ et K+ en favorisant un nombre de coordination élevé et une géométrie irrégulière. Avec un nombre de charge de +2, Ca2+ peut être lié à des anions que les ions alcalins ne peuvent pas ;
c) Les vitesses de fixation et de dissociation sur Ca2+ sont élevées : la vitesse de fixation est limitée par la diffusion et la vitesse de dissociation correspond à la stabilité. La vitesse d’exchange sur Mg2+ est
105s-1, plus lente que celle Ca2+de presque trois ordres de grandeur. Cette vitesse d’exchange rapide, qui est celle d’un système très réactif, explique l’utilisation du Ca2+ quand des réponses rapides sont nécessaires comme l’ouverture et la fermeture des canaux membranaires, pour la régulation du transport ionique et la contraction musculaire.
Les protéines qui fixent le calcium sont en général riches en aspartate et en glutamate, qui portent tous deux des groupements carboxylate sur leur chaîne latérale et qui peuvent donc agir comme ligands anioniques durs. Comme illustré dans une forme de structure qui existe, un Ca2+ entouré de 4 carboxylate avec des oxygènes donneurs, la géométrie est un tétraèdre déformé (figure 2). La structure a été caractérisée par diffraction des rayons X.
L’ion Ca2+ fonctionne communément comme un pont entre différents segments de la protéine. Il est lié aux groupements latéraux anioniques d’aminoacides différents, ou même à des groupements carbonyle (figure 2). L’utilisation de cette propriété dans le contrôle du repliement de la chaine est illustrée schématiquement sur la figure 2 qui montre comment la chaine peptidique se replie pour permettre aux quatre COO- de se coordiner au Ca2+. Le résultat de ce mouvement est le déplacement des emplacements des groupements latéraux de l’une des régions hélicoïdales de la protéine par rapport aux groupements latéraux de la deuxième région hélicoïdale.
Les variations du repliement des protéines contrôlent la structure et les fonctions de la cellule telles que sa vitesse de croissance et son métabolisme énergétique.
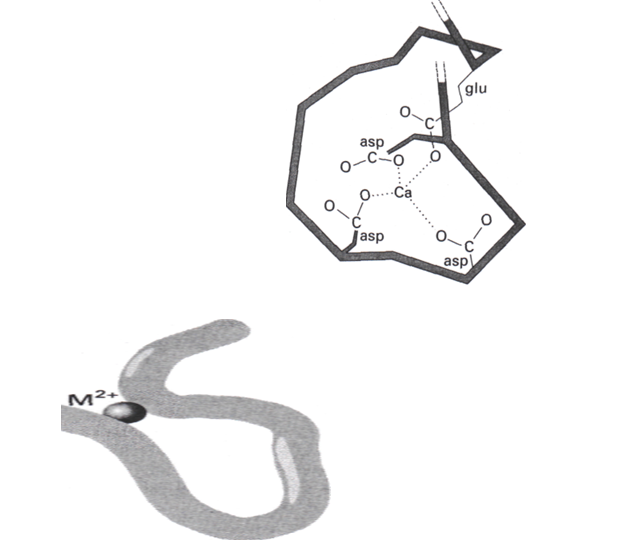
Figure 2
La position des ions Ca2+ et Mg2+ influence les déformations des protéines et par conséquent change les structures et fonctions des cellules, la labilité du Ca2+ permet les changements structuraux rapides.